隐花色素光反应机制新发现
隐花色素为自然界最广泛存在的光受体之一,于上世纪在拟南芥中率先发现,后陆续被证实存在于包括人类在内的所有动植物中。隐花色素具有许多生物学功能,比如调控植物生长发育、调控人体生物钟与多种疾病的发生等。近日,福建农林大学基础林学与蛋白质组学研究中心王琴课题组在Molecular Plant杂志在线发表了题为“Photooligomerization determines photosensitivity and photoreactivity of plant cryptochromes”的研究论文(https://doi.org/10.1016/j.molp.2020.01.002),报道了植物隐花色素早期光反应机制的新发现。柳青博士、苏甜甜博士和何文锦博士为本文共同第一作者,王琴教授为本文通讯作者。

在该文中,作者对隐花色素的早期光反应即蓝光所催化的光受体蛋白多聚化反应首次进行了光动力学定量研究。该研究分析了多个有关该光受体的有趣现象,回答了一些动植物光与时间生物学上的新老问题。首先,隐花色素的多聚化反应到底有多快?是否可逆?作者发现隐花色素的多聚化反应极为迅速,例如植物细胞中的拟南芥隐花色素2(CRY2)蛋白可在接收蓝光后数十秒内完成多聚化反应。进入黑暗后,多聚化的隐花色素蛋白又能发生逆转而自发恢复成单体形态,而该逆反应的速度相对于光反应则慢了不少,CRY2在植物细胞
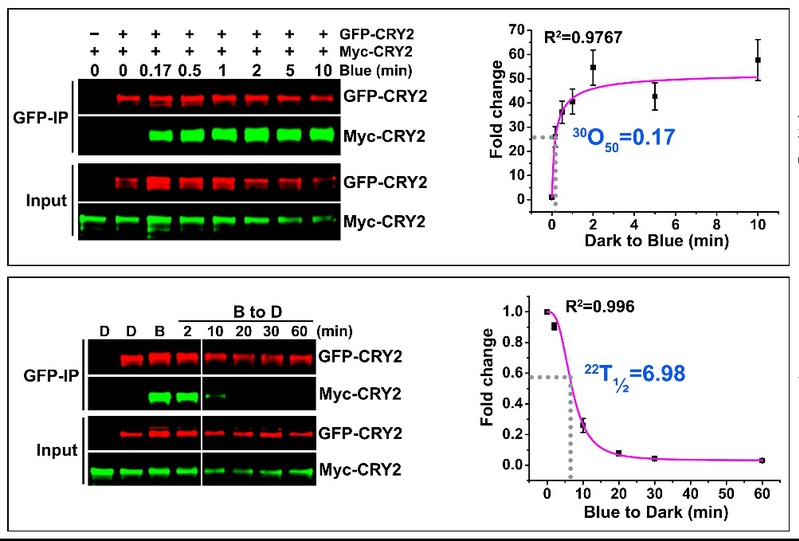
中由多聚态到单体态的反应需要数分钟。
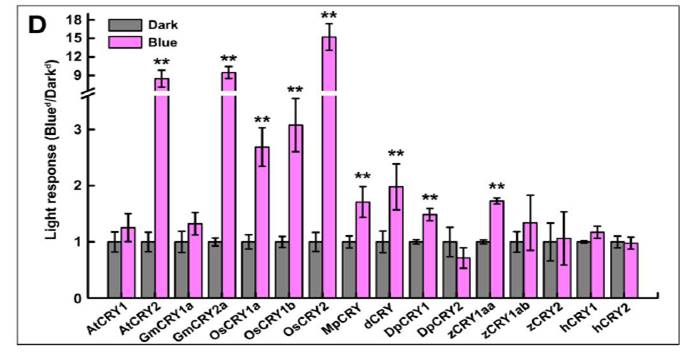
其次,虽然隐花色素的蛋白多聚化反应已在多年前被中国、德国与美国的多个实验室证实为其生理生化功能所需的必要条件,但该反应是否为隐花色素生理生化功能的充分条件这一问题则多年悬而未决。就此,作者筛选了数个CRY2点突变体并对其光多聚化特性与生理生化功能进行了分析。作者发现隐花色素的多聚化是其功能的必要但非充分条件。由此推测隐花色素多聚化后可能还存在进一步构象变化以改变隐花色素与其它蛋白的相互作用从而发挥其正常的生理生化功能。作为一类古老的光受体,其任何生化反应都存在着一个进化上保守度的问题。例如哪些隐花色素具有蓝光催化的多聚化反应?解决该问题有助于我们了解这一类光受体的进化历史,以便人们能够更好地了解并在农业与医学上利用这些光受体。作者发现隐花色素光多聚化是一种进化上极为保守的现象,并与生物体中隐花色素的光反应特性高度相关。例如大部分植物隐花色素均为光受体而它们也都能够发生蓝光催化的多聚化反应,而动物隐花色素只有曾经报道过具有光反应活性的隐花色素能够发生蓝光催化的多聚化反应(如果蝇dCRY,帝王蝶DpCRY1和斑马鱼zCRY1aa)。该实验结果的广泛生物学含义还有待于进一步的研究与论证。
最后,大部分生物体内都不止一个隐花色素。例如拟南芥与人有均分别有两个隐花色素(CRY1,CRY2),水稻有三个隐花色素(OsCRY1a,OsCRY1b,OsCRY2),而大豆则有七个隐花色素(GmCRY1a-d,GmCRY2a-c)。以往的研究发现不同隐花色素有着相似以及独特的功能。但是同一细胞中不同隐花色素功能之间有何关联尚不清楚。作者发现同一植物的不同隐花色素能发生异源光多聚化反应。例如拟南芥的CRY1与CRY2不仅在蓝光催化下可完成自身的CRY1-CRY1与CRY2-CRY2同源多聚化,同时还能形成CRY1-CRY2异源多聚化。为了解隐花色素异源多聚化现象的潜在生物学功能,作者将拟南芥CRY2的截短蛋白(即N端PHR2)过表达到野生型植物背景中,结果显示这些植物在蓝光下呈现出了长下胚轴的表型,这属于典型的cry1突变体表型。为什么表达了大量PHR2蛋白的植物失去了CRY1的功能呢?进一步的实验表明细胞内大量积累的PHR2蛋白能够通过异源多聚化反应与细胞内的CRY1与CRY2相互结合,从而抑制了CRY1的同源多聚化,进而抑制CRY1的功能,导致PHR2过表达植物的下胚轴伸长生长在蓝光下无法受到蓝光的抑制,这个结果也表明蓝光不仅能催化隐花色素同源多聚化使这些光受体行使各自的功能,同时还可能通过催化不同隐花色素间的异源多聚化以达到调节细胞内不同隐花色素行使各自独特功能的目的。
阅读原文: https://doi.org/10.1016/j.molp.2020.01.002










